Overview Chromosomes (Chromosomae):
Pages with explanations are linked to the
text below the images if available! (Labelling is in German)
 |
 |
 |
 |
 |
 |
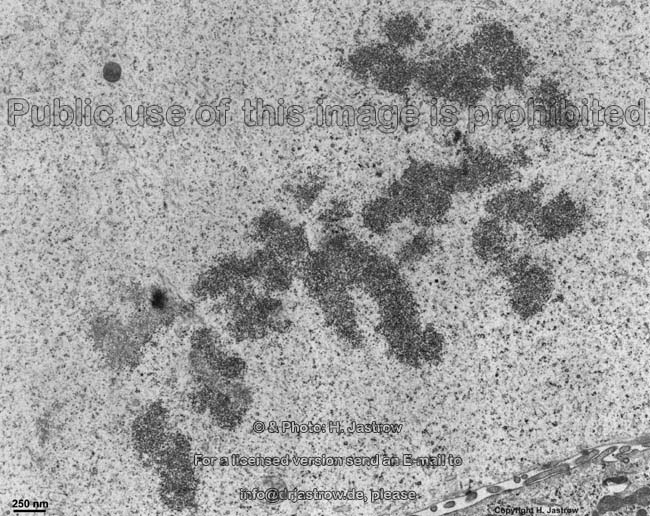
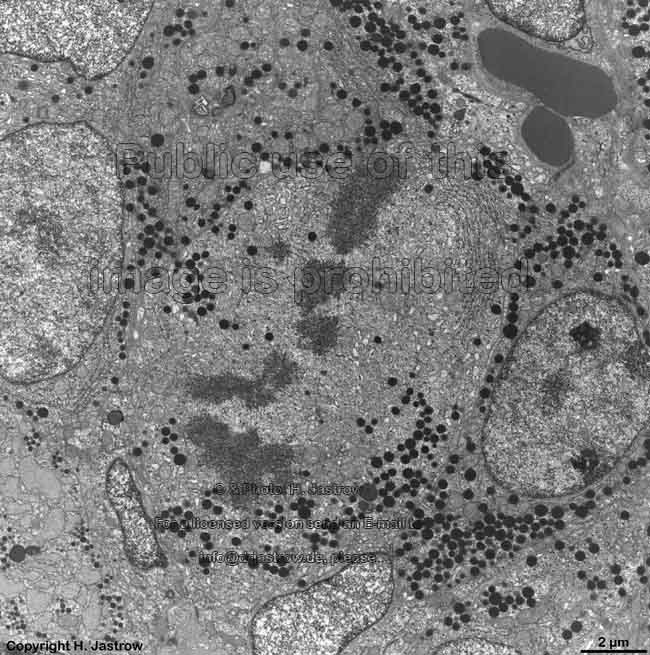
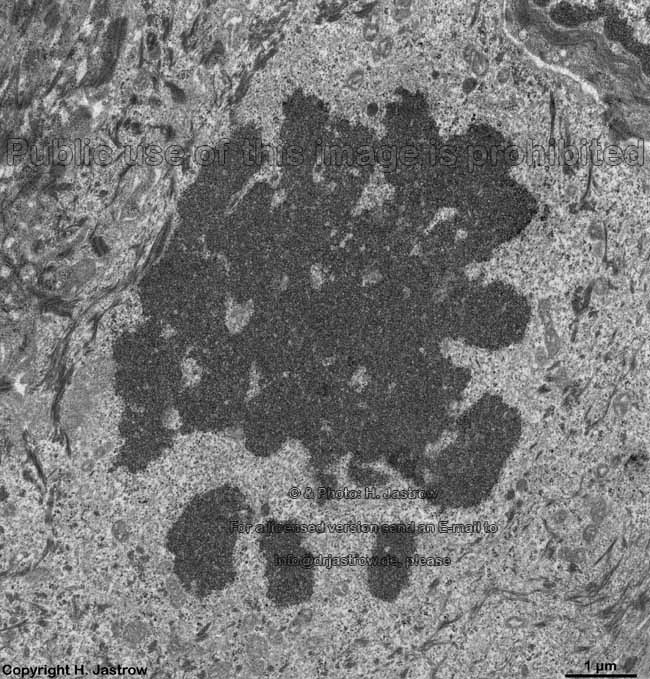
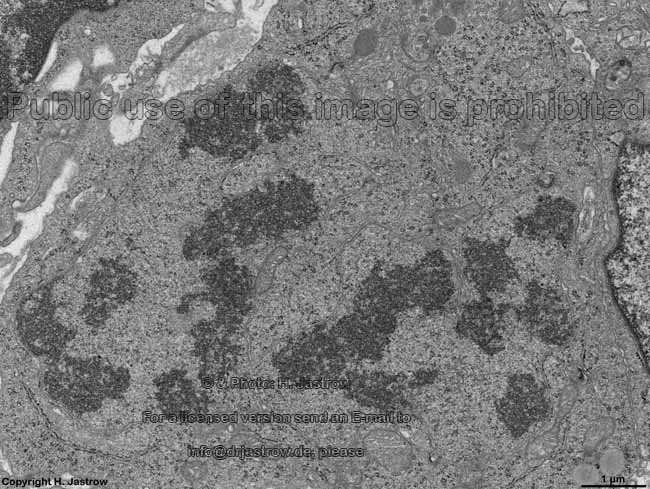
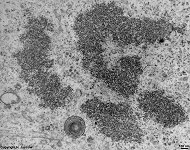
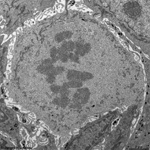
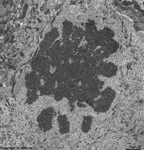
chromosomes in
metaphase (rat) |
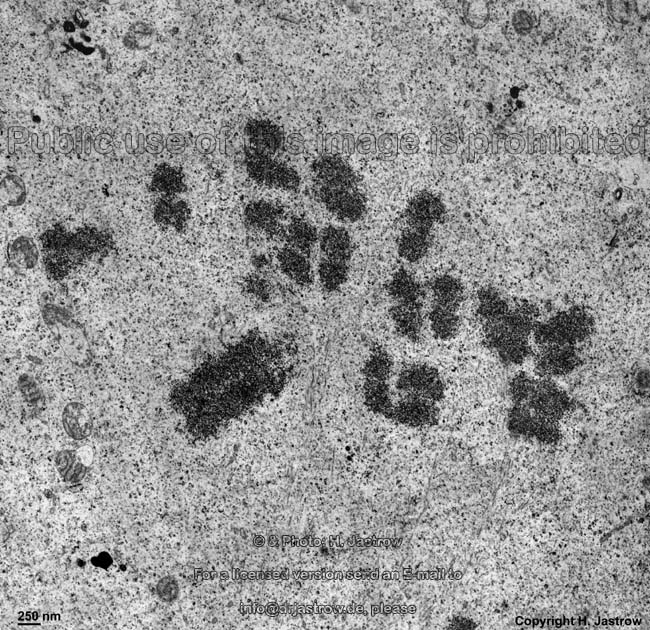
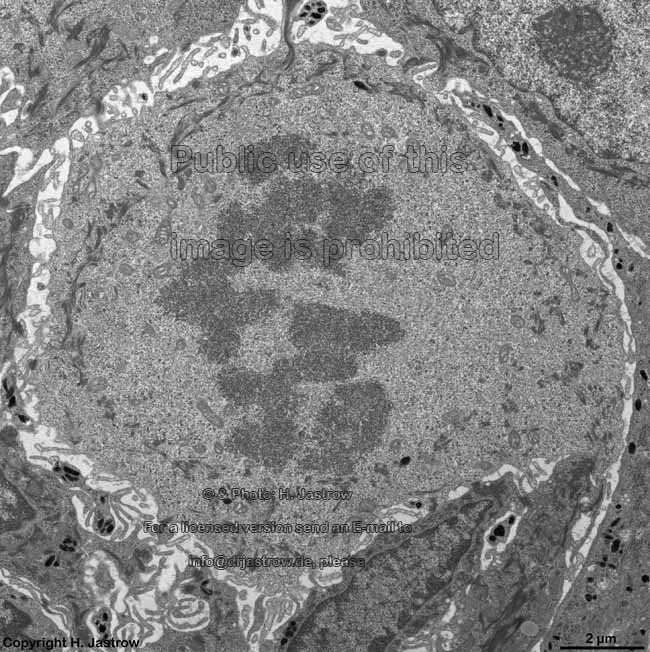
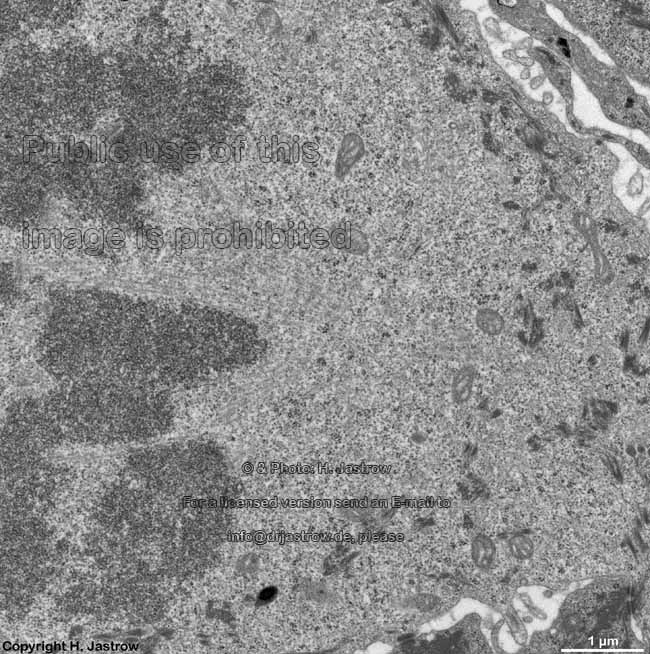
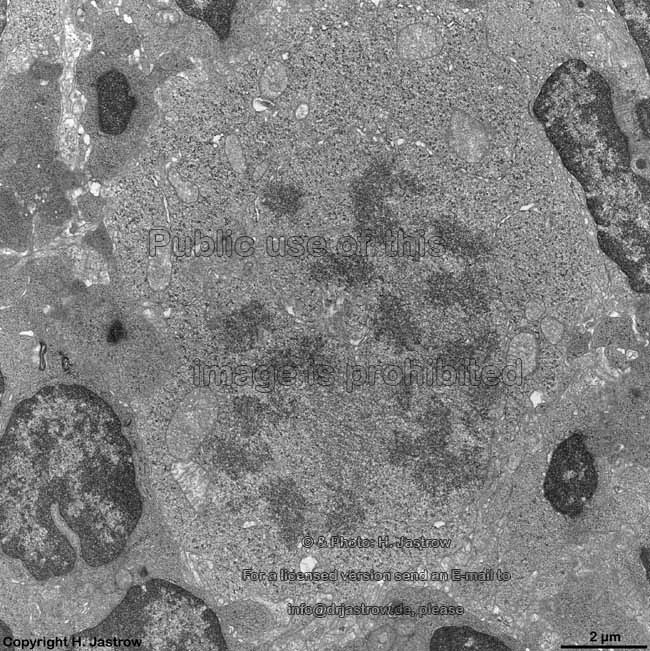
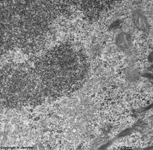
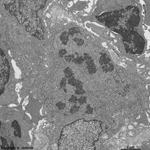
chromosomes
Claudius cell (rat) |
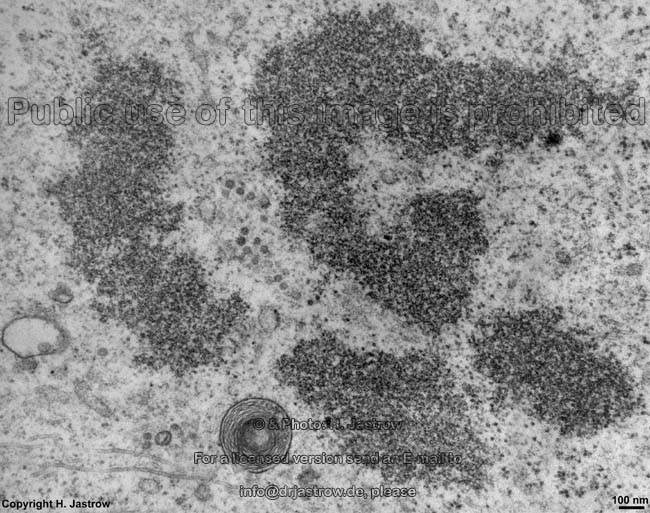
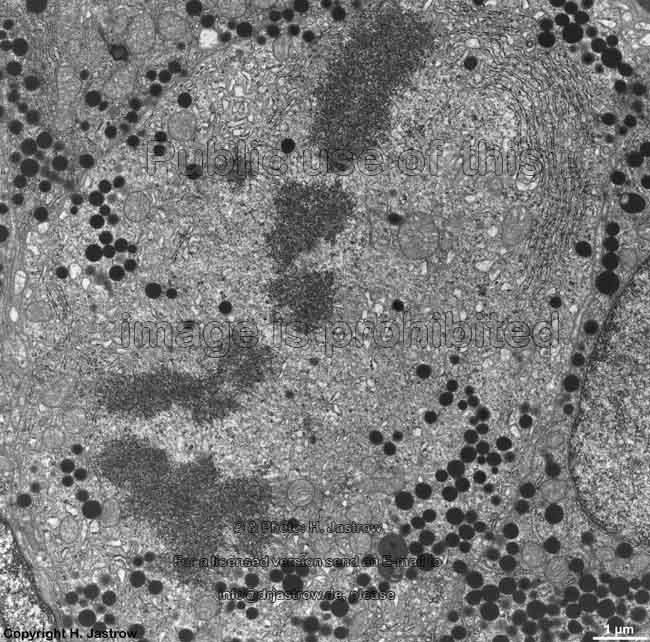
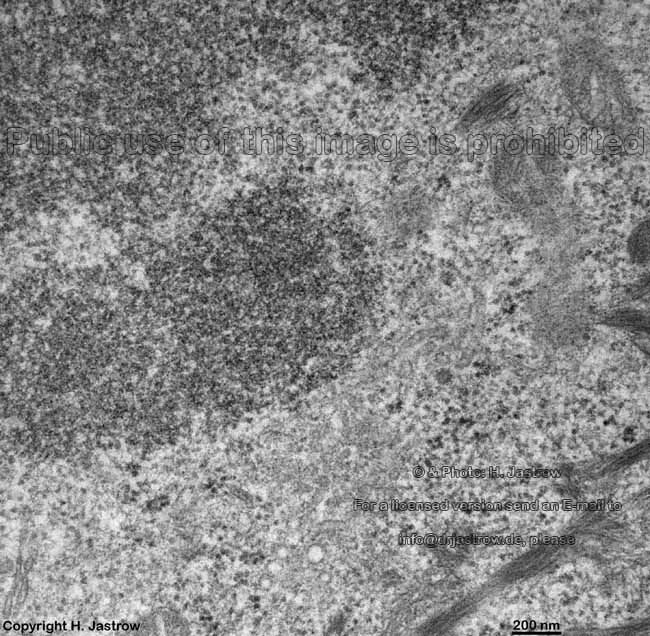
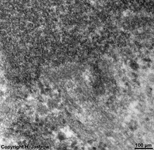
chromosom
detail (rat) |
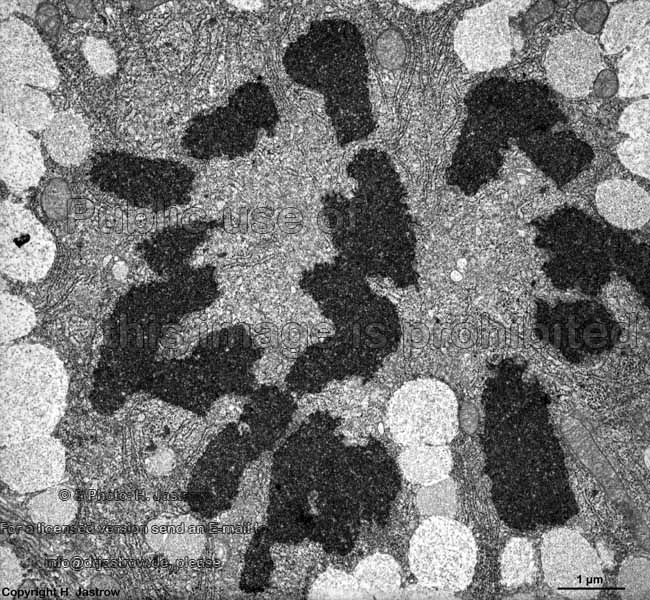
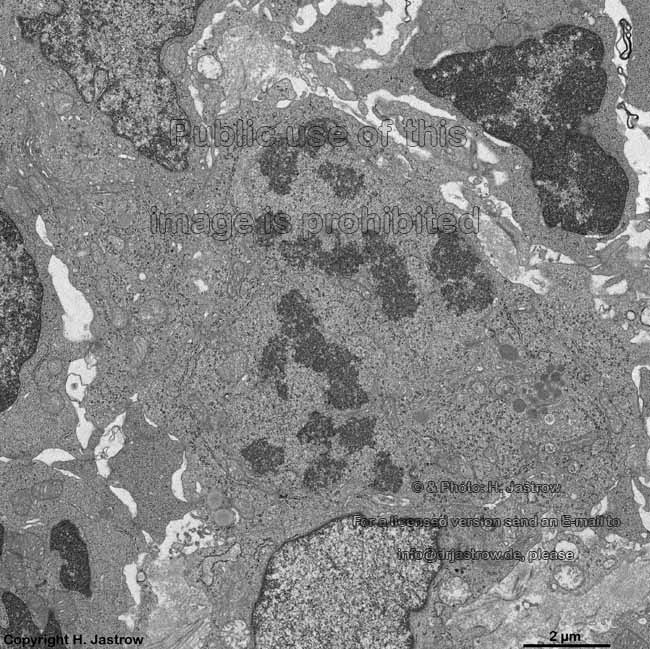
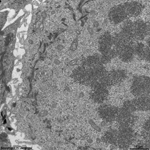
chromosomes + multi-
lamellar body (rat) |
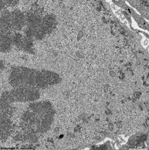
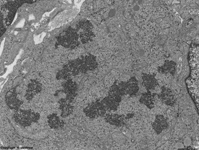
chromosoms of a seba-
ceous gland cell (rat) |
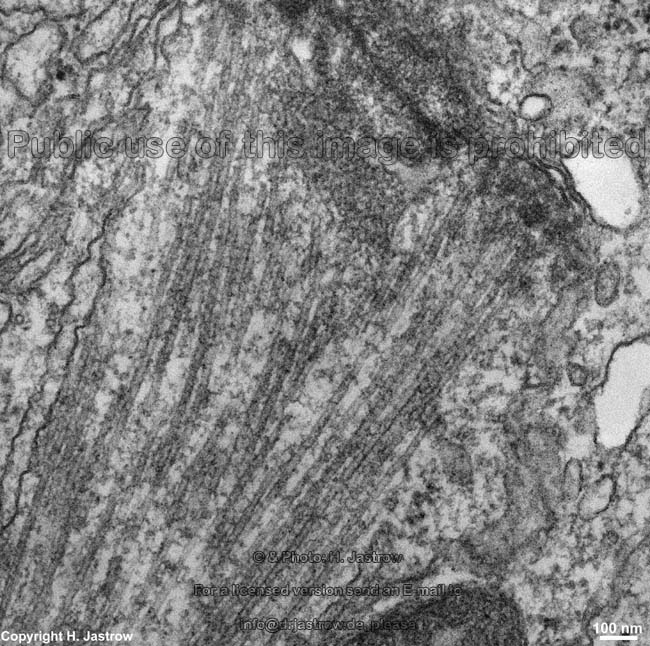
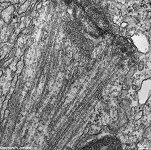
Kinetochor +
spindle fibres (rat) |
|
|
|
|
|
|
|
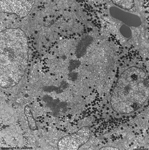
chromosomes
hypophsis (rat) |
detail: pairing in
metaphase |
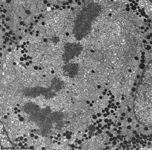
skin (human)
anaphase |
detail 1 |
detail 2 |
detail 3 |
human skin
prometaphase |
|
|

|

|

|

|
|
Tonsilla pha-
ryngea (human) |
detail: formation of chro-
mosomes in prophase |
Tonsilla pharyngea
Prophase 2 (Mensch) |
detail 1: chromosome
formation |
detail 2 |
detail 3 |
Kinetochor liver
cell (rat) |
Chromosomes (Terminologia histologica: Chromosomae) are the transport
form of genetic information during cell division, i.e. mitosis
or meiosis. They are formed by condensation and helical conformation of
desoxyribonucleic acid (DNA) which carries the
genetic infomation. DNA consists of 2 hlically arranged chains of molecules
of desoxyribose, a sugar molecule interconnected by phosphate groups. One
of the four nucleotids (Adenin [A], Cytosin [C],
Guanin [G] or Thymin [T]) is connected to one carbon nucleus
of each sugar molecule. The basic nucleotids of one chain tightly connect
to those of the other chain forming the base pairs. Hereby A
in chain 1 is always coupled to T of chain 2 and C of chain
1 is tighted to G of chain 2 and vice versa. The length of the longest
human DNA chains comprised of ~250.000.000 base pairs, is about 10 cm,
the diameter only 2 nanometers (nm; = 0.0000002 cm). Each chromosome has
only one linear DNA molecule. For a reasonable spatial arrangement the
DNA is wired around histone proteins which are also components of chromosomes.
There are 5 major types of histon proteins (H1, H2A, H2B, H3 and
H4). Histone proteins 2-4 arrange to disk-like octamers, i.e. units
of 8 linked proteins consiting of 2 molecules of H2A, H2B, H3 and H4 each.
Since these proteins are rich in basic amino acids the negatively charged
phosphate groups of the DNA bind to them. Hereby the DNA winds around the
histone octamers in a manner that about 146 base pairs are wired around
each octamer. This unit of histone octamer and attached base pairs is called
nucleosome
and has a diameter of ~11 nm. Thus the DNA appears like a chain of pearls
whereby the spheres are formed by the nucleosomes interconnected by thin
sections of the DNA stripe. The chains of pearls are called chromatin
fibrils. The formation of the nucleosomes is supposed to be managed
by 2 additional proteins in presence of DNA: nucleoplasmin and N1 protein,
that are not inculded into the octamer and thus called non-histone proteins.
The histone protein 1 H1 which is not included in nucleosome formation
attaches to the DNA opposite to the nucleosomes and links with neighbouring
H1 units to form a framework causing a larger helical arragngement of nucleosomes.
Further scaffold proteins attach to the H1 e.g., the enzyme topoisomerase
II. The resulting tubes of arranged chromatin fibrils have diameters of
about 30 nm. The 700 nm wide chromosome sections are formed by further
folding of the tubes and winding around each other. They are densely packed
in form of the chromatids. Both chromatids of a chromosome are linked
to each other at theCentromer (= primary
incisure). Specific sequences of nucleic acids present in the centromer
called CEN sequence. The centromer-binding factor, a complex of 3 proteins,
binds to this sequence of the DNA. It on the other hand has binding sites
for microtubules that attach here during
mitosis.
These microtubules are also called spindle
fibres and attach to opposite sites of the centromer at the daughter chromatids
to cause their separation. A kinetochor
consists of disk-like stacks of protein layers (inner, central and outer)
in the centromer region of a chromosome. A diffuse corona attaches to the
outer lamella which contains the microtubule-binding protein (CLIP-170)
like the outer lamella and the microtubular motor proteins dynein and CENP-E.
The kinetochor binds to the plus end of microtubules.
The number of chromosomes of organisms is species specific and constant.
With exception of the ova and spermatocytes
all other somatic cells of humans have, with exception of few multiploid
cells, a double set of chromosomes, i.e. they are diploid comprised of
46 chromosomes, 44 autosomes and 2 gonosomes. Autosomes are
all chromosomes bearing the genetic information for all non-sex specific
protiens of the body. One of the 2 chromosomes in each pair is from the
mother, the other from the father. These corresponding chromosomes that
carry the same genes are called homologous chromosomes. The gonosomes
which are also called heterosomes or sey chromosomes carry a large number
of genes with the information of sex-specific proteins and of proteins
required for the regulation and formation of primary and secondary sex
organs. There are 2 different gonosomes, the X-chromosome and the Y-chromosome.
While all cells in females have a pair of X-chromosomes,
cells of males show 1 X- and 1 Y-chromosome. In females
only the inforation of one X-chromosome is used whereas the second X-chromosome
is inactive and condensed as a X-chromatin or heterochromatic body (Barr
body). Such a Barr body is detectable in cells of the oropharyngeal
mucosa with the light- or electron microscope as a lump of heterochromatine
which has a diameter of 1 - 2 µm and which is attached to the inner
membrane of the cell nucleus.
The female reproductive cells (ova) are
haploid, i.e. posses only a half set of chromosomes in which all autosomes
are only present once plus one gonosome which always is an X-chromosome
(number of chromosomes: 22 + X). Male reproductive cells, i.e. spermatocytes
are also haploid with the difference that their gonosome may either be
an X- or an Y-chromosome. By fertilisation the ovum
gets the chromosomes of the spermatozoon
and the resulted fertilised ovum has a full (diploid) set of chromosomes
(either femal: 44 + XX or male: 44 + XY). The vast majority of cells is
euploid,
i.e. diploid with complete set of 46 chromosomes in the
cell
nucleus. In some organs, however, certain cells may have multiple sets
of chromosomes. This means they are polyploid. An example of polyploid
cells are hepatocytes. It is assumed that this is caused by the extremely
high metabolic activity. Usually these cells are tetraploid, i.e. have
a 4x set of chromosomes. Occasionally, hexaploid (6x) or octaploid (8x)
cells are encountered. In contrast to that tumor cells may be triploid
(3x) or pentaploid (5x) or other irregular sets of chromosomes.
The shape and the size as well as the bands of chromosomes in light
microscopic staining are constant allowing a classification using the size
and length of chromosome branches to the centromer. In a karyogram
all chromosomes are arranged according to their size using a micrograph
of usually a lymphocyte in light microsopic stain
after administration of a drug which stops mitosis in metaphase
when chromosomes are best visible. The best stain for visualisation of
the bands of chromosomes is orcein. The visible bands are caused by the
differences of chromatin fibil density in the regions of the chromosomes.
Further the relation of A-T to G-C pairs has an influence on stain. The
typical bands with size and location of the centromere allow to create
karyograms which are important for genetic investigations to detect abnormalities
e.g., trisomy (one chromosome is present 3x) or monosomy (present only
once). The micrographed chromosomes have 2 long processes, the chromatids
which are connected at the centromer as mentioned above. According to the
position of the centromer we can distinguish: metacentrisc chromosomes
(centromer in the centre: chromosomes 1,2,3,11,19,20,X), submetacentric
chromosomes (centromer not exactly in the centre: chromosomes 6,7,8,12),
acrocentric
chromosomes (centromer located close to the end: chromosomes 13,14,15,21,22,Y),
subacrocentric
chromosomes (centromer located closer to the end than to the centre: chromosomes
4,5,9,10,16,17,18). Since even in metacentric chromosomes the centre is
not absolutely exactly in the middle it is possible to always distinguish
2 longer and 2shorter arms of each chromosome. The longer ones are labelled
with q and the shorter ones with p. Further there are secondary
laces in the short arms of the acrocentric chromosomes 13, 14, 15,
21 and 22. These locations are the nucleolus-organisator centres
which are responsible for the formation of nucleoli.
The ends of the short arms of these chromosomes are called satellits
behind the incisure.
Chromosomes are electron-dense structures in a transmission
electron microscope which can only be seen in prometaphase,
metaphase
and anaphase of the mitosis. They are 2 to
10 µm in length and both chromatids are about 0.5 µm in thickness.
A clear determination which chromosome is regarded is not possible without
special procedures.
--> Euchromatine, heterochromatine,
synaptonemal
complex, cell nucleus, mitosis
--> Electron microscopic atlas Overview
--> Homepage of the workshop
Images, page & copyright H. Jastrow.